I - Intéraction Hormone/Récepteur:
Les hormones se fixent sur des récepteurs spécifiques et induisent l'activation ou l'hinibition des cellules cibles (réponse cellulaire).
On distingue 4 types de récepteurs:
- R. couplés à une protéine G
- R. couplés à une réaction enzymatique
- R. couplés à un canal ionique ou étant des canaux ioniques
- R. assossiés à l'ADN (intéraction avec un promoteur)
II - Les récepteurs = protéines de liaison spécifique:
Les récepteurs ont une spécificité relative.
Les récepteurs ont une forte affinité (Kd = 10-9 à 10-12).
Les récepteurs ont un nombre de sites limités.
Les récepteurs sont spécifiques de l'expression tissulaire (épissage alternatif).
La liaison hormone/récepteur est réversible:
Les liaisons S (spécifiques) et NS (non spécifiques) sont réversibles. Le plateau (NS) du graph 2 sont les liaisons non spécifiques.
II.1 -Régulation des récepteurs hormonaux:
- Modification qualitative:
Baisse de l'affinité due à une acidose ou due à une phosphorylation par une Protéine Kinase.
Masquage des sites, quand plusieurs sites sont occupés il y a une baisse de l'affinité.
- Modification quantitative:
(1) Sur la membrane, il y a fixation d'une hormone sur un récepteur.
(2) Migration du complexe et formation d'une couverture de clatherine.
(3) Endocytose de la vésicule.
(4) Transport au niveau de l'appareil de Golgi et association (5).
Deux destinations sont possibles :
(6) Dégradation par protéolyse lysosomiale du récepteur et de l'hormone.
(7) Recyclage du récepteur sur la membrane.
=> Diminution de la capacité de réponse de la cellule quand il y a une trop forte stimulation hormonale.
II.2 - Régulation par l'hormone:
- Up régulation : la synthèse d'hormone induit une augmentation de l'expression génétique et la synthèse de récepteurs.
- Down régulation : plus il y a d'hormone moins il y a de récepteurs (diminution de l'expression génètique codant pour le récepteur et phénomène d'internalisation).
III - Récepteurs membranaires couplés aux protéines G:
Ces récepteurs sont membranaires, associés à des protéines G, elles mêmes à proximité d'une Adénylate cyclase.
Ces récepteurs ont :
- 7 domaines transmembranaires.
- Une partie N terminale extracellulaire où se fixe l'hormone.
- Une partie C terminale intracelllaire où il y a des sites de phosphorylation qui modulent l'affinité et une boucle nécessaire à la fixation des protéines G.
III.1 - Protéine G : couplage récepteur et transduction intracellulaire:
L'activation des protéines G via le récepteur à hormone conduit à la transduction d'un signal intracellulaire qui peut être de 2 natures : via l'AMPc ou via le Ca2+.
L'AMPc (adénosine mono phosphate cyclique) active les protéines kinases AMPc dépendantes = PKA.
Le Ca2+ (ion calcium) active les protéines kinases Ca2+ dépendantes = PKC.
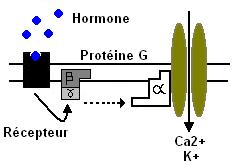
Les protéines G sont constituées de 3 sous unités : alpha, béta et gamma.
La sous unité alpha peut être sous 2 formes S ou I.
Alpha S est stimulatrice, se lie au GTP, l'hydrolyse et ainsi active une enzyme membranaire, l'adénylate cyclase.
Alpha I est inhibitrice, se lie au GTP, l'hydrolyse afin d'inhiber l'adénylate cyclase.
Béta et gamma associent alpha à la membrane et peuvent stimuler des PKinases.
III.2 - Activation des potéines G
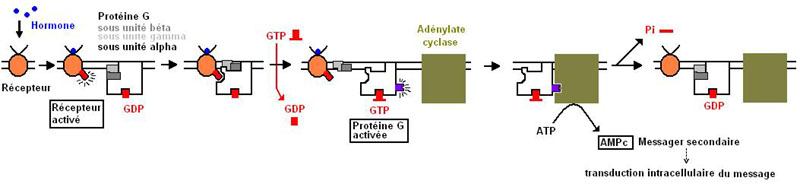
Au repos la Ptn G (sous unité alpha) a une forte affinité pour le GDP (Guanosine di-phosphate).
Quand l'hormone se fixe au récepteur, ce dernier change de conformation et laisse apparaître un site de fixation
pour la Ptn G. La Ptn G migre et se fixe. Sa structre est modifiée et son affinité pour le GDP diminue au dépend du GTP.
Le GTP se fixe induisant l' apparition d'un site de fixation pour l'adénylate cyclase et la dissociation de la sous unité alpha des deux autres. La sous unité alpha "active" migre jusqu'à l'Adénylate cyclase et se fixe.
De l'ATP intracellulaire est transfomé en AMPc, messager secondaire intracellulaire.
L'hydrolyse du GTP en GDP fait disparaitre le site de fixation et la sous unité alpha se réassocie aux deux autres sous unités.
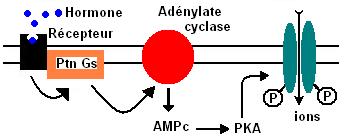
III.2.1 - Action hormonal via la voie AMPc :
La concentration d'AMPc intracellulaire dépend de la synthèse et la dégradation d'une phosphodiestérase.
L'AMPc active les protéines kinases AMPc dépendantes (PKA).
La PKA est constitué de 4 sous-unités, 2 régulatrices (fixation AMPc) et 2 catalytiques (phosphorylation).
La PKA est inactive, il faut 2 molécules d'AMPc pour l'activer; les deux sous-unités catalytiques se dissocient et peuvent phosphoryler d'autres protéines.
Exemple du glucagon :

L'AMPc peut intervenir sur la régulation de l'expression des gènes par la phosphorylation de la protéine CREB (CAMP response element binding protein) via la PKA. La protéine CREB se dimérise et se fixe sur un promoteur ADN spécifique (CRE) avec in fine production de protéines.
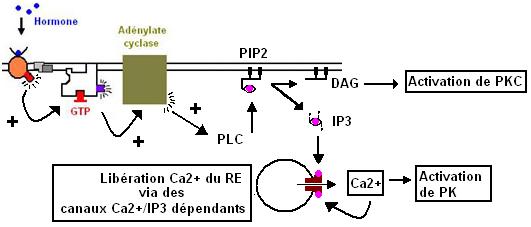
III.2.2 - Action hormonale via la voie Ca2+ :
Dans cette voie il y a intervention d'un phospholipide membranaire: le phosphatidylinositol-bisphosphate ou PIP2.
PIP2 est hydrolysé par une phospholypase de type C en Diacylglycérol (DAG) et Inositol trisphosphate (IP3).

Il y a couplage récepteur/Ptn G qui par l'adénylate cyclase active la PLC.Le Ca2+ est soit libéré du RE soit entre dans la cellule. Ca2+ s'autoactive en se fixant sur les canaux. IP3 est dégradé par une phosphatase arrêtant ainsi la libération de Ca2+. Au repos, quand il n'y a pas de stimulation hormonal, le Ca2+ est expulsé de la cellule par des canaux Ca2+/ATPase et des canaux antiports Na+/Ca2+ dependant.
Le RE et les mitochondries possédent aussi des système de pompage du Ca2+.
Dans le cytoplasme il y a des molécules qui captent la calcium, les CADMODULINES.
Au repos, la concentration intracellulaire de Ca2+ est faible (10
-7M). Une petite entrée de calcium provoque une grande variation de [Ca2+]i et active le mécanisme de réponse cellulaire.
Le Ca2+ se fixe à la cadmoduline. Le complexe formé se fixe et active une protéine Kinase Ca2+/Cadmoduline dépendante, la CAM Kinase.
La fixation permet aussi la libération d'un site de phosphorylation sur lequel la CAMK activée peut s'autophosphoryler. La CAMK est ainsi totalement active.
Si la concentration intracellulaire - [Ca2+]i - diminue, le grand complexe CAMK/Cadmoduline/Ca2+ se dissocie mais la CAMK reste active tant qu'elle est phosphorylée. L'activation est indépendante de Ca2+.
L'inactivation de la CAMK se fait par une phosphatase.
La CAMK active a pour rôle de phosphoryler des enzymes et des protéines activatrices et d'activer l'expression
génètique (phosphorylation de MAP kinases qui vont phosphoryler des protéines inhibitrices de transcrption fixées aux gènes. Phosphorylation de facteurs de transcription).
Action des hormones via l'acide arachidonique :
Le récepteur d'hormone est couplé à une protéine G. La stimulation de cette Pt G induit l'activation de la phospholypase A2 (PLA2)) qui hydrolyse les phospholipides membranaires.
La PLA2 coupe un groupement acyl (l'acide arachidonique) au niveau d'un phosphatidyl-(A) et laisse dans la membrane un lysophospholipide.

Le phosphatidyl-(A) peut être le phosphatidyl - choline,sérine, éthanolamine ou inositol.
L'acide arachidonique peut activer des enzymes cellulaires (via une PKC), et peut être utilisé comme métabolite pour la synthèse de Prostanoïdes, Leucotriènes (médiateur paracrine) ou d'époxydes activation de protéines GMPc dépendantes).
Récepteurs à activité enzymatique :
Récepteurs ayant un domaine extracellulaire et un domaine intracellulaire et comportant une activité enzymatique intrinséque ou à association directe.
Récepteurs à activité guanylate cyclase (GMPc) :
Exemple de l'ANF (facteur atrial natriurétique) :
L'ANF agit sur le coeur en régulant la pression artérielle et sur l'équilibre hydro-minéral. Le récepteur activé par l'ANF synthètise du GMPc. Le GMPc active les protèines Kinases G qui vont phosphoryler d'autres protéines. Les PKG sont généralement des Serine kinases ou des thréonine Kinases.
Récepteurs à activité Tyrosine Kinase :
Ces récepteurs sont pour la plupart des facteurs de croissance. Ils ont une structure transmembranaire comportant un domaine extracellulaire (beaucoup de cystéines et de ponts dissulfures et fixation de l'hormone) et un domaine intracellulaire (activité tyrosine kinase).

Sans hormone il n'y a pas d'activation du recepteur.
Quand l'hormone se fixe, le récepteur est actif, il peut alors se dimèriser et s'autophosphoryler pour être totalement activé.
La phosphorylation s'effectue sur des résidus Tyrosine.
Cela mène à l'activation d'une GTPase qui est à la base d'une activation séquentielle de protéines jusqu'à celle
d'une MAP Kinase. MAP K est activée pendant l'activité mitogène d'une cellule.
Récepteurs associés aux tyrosines kinases :
Récepteurs de médiateurs locaux produits par le sytème immunitaire (les cytokines). Ces récepteurs sont constitués de plusieurs sous-unités qui s'associent lorsqu'une hormone se fixe.Ce récepteur actif s'associe et active une protéine tyrosine kinase de type Src ou Janus Kinase. (c'est par exemple le cas des récepteurs à IL2).
Les récepteurs intracellulaires
Ils ont tous une structures voisine et appartiennent à une seule et même famille de récepteurs.
On trouve par exemple les récepteurs aux stéroïdes et aux hormones thyroïdiennes.
Ces récepteurs sont des protéines activatrices de l'expression génètique. Ils se fixent sur des régions promotrices (HRE : hormon response element) de gènes sous forme de dimères. Ces dimères sont des homo ou des hétéro-dimères (exemple des protéines à doigt de zinc)
Récepteurs associés aux protéines HSP90 :

Récepteurs aux stéroïdes :
Inactif avant la liaison avec l'hormone et comportant HSP90 sur le site de liaison à l'ADN.
Quand il y a fixation de l'hormone, il y a un changement de conformation du récepteur, libérant HSP90 et laissant le site de liaison à l'ADN libre
et actif. Le complexe migre vers le noyau où il intéragit avec les séquences promotrices spécifiques de gènes cibles.
Récepteurs liés à l'ADN :
C'est le cas des récepteurs des hormones thyroïdiennes. Le récepteur est déjà sur la séquence HRE (TRE pour les ho thyroïdiennes) du gène cible.
L'hormone T3 est transportée dans le sang (par une protéine de transport) puis dans le cytoplasme (par une protéine de transport) et jusqu'au noyau pour sa fixation sur son récepteur.
Le complexe Ho/R active des facteurs de transcription (Ft et Pol2) qui activent la polymérase II et ainsi débute l'expression du gène. Les récepteurs nucléaires sont des facteurs transcriptionnels ligand dépendant.
Les récepteurs canaux ou liés à des canaux
Récepteur nicotinique à l'ACTH
composé de 5 sous-unités : 2 alpha, 1béta, 1 gamma et 1 delta. Elles forment un canal
Récepteurs couplés à un canal par phosphorylation
Récepteurs couplés à un canal par une protéine G